Формирование хромосом происходит в момент мейоза, когда в процессе кроссинговера, каждая случайно берет примерно половину от материнской хромосомы и половину от отцовской, какие конкретно гены достанутся от мамы, а какие от папы - не известно, все решает случай.
В этой лотереи не участвует только одна мужская хромосома - Y, она целиком передается от отца к сыну как эстафетная палочка. Уточню, что у женщин данной Y-хромосомы нет совсем.
В каждом последующем поколении в определенных участках Y-хромосомы, называемых локусами, происходят мутации, которые будут передаваться всем последующим поколениям по мужскому роду.
Именно благодаря этим мутациям стало возможным реконструировать рода. На Y хромосоме всего примерно 400 локусов, но для сравнительного анализа гаплотипов и реконструкции родов используется только около сотни.
В так называемых локусах, или их еще называют STR-маркеры, бывает от 7 до 42 тандемных повторов, общая картина которых является уникальной для каждого человека. Через определенное количество поколений происходят мутации и количество тандемных повторов изменяется в большую или меньшую сторону и таким образом на общем древе будет видно, что чем больше мутаций, тем древнее общий предок для группы гаплотипов.
Сами гаплогруппы не несут генетической информации, т.к. генетическая информация находиться в аутосомах- первых 22 пар хромосом. Посмотреть распространение генетических компонент в Европе можно . Гаплогруппы являются всего лишь метками давно минувших дней, на заре формирования современных народов.
Какие гаплогруппы у русских наиболее часто встречаются?
Народы |
человек |
||||||||
|---|---|---|---|---|---|---|---|---|---|
Восточные, западные и южные славяне . |
|||||||||
| Русские (север) | 395 | 34 | 6 | 10 | 8 | 35 | 2 | 1 | |
| Русские (центр) | 388 | 52 | 8 | 5 | 10 | 16 | 4 | 1 | |
| Русские (юг) | 424 | 50 | 4 | 4 | 16 | 10 | 5 | 3 | |
| Русские (все великороссы) | 1207 | 47 | 7 | 5 | 12 | 20 | 4 | 3 | 2 |
| Белорусы | 574 | 52 | 10 | 3 | 16 | 10 | 3 | Русские, славяне, индо-европейцы и гаплогруппы R1a, R1b, N1c, I1 и I2В глубокой древности, около 8-9 тысячелетий назад была лингвистическая группа, положившая начало индо-европейской семье языков (на начальном этапе вероятнее всего это гаплогруппы R1a и R1b). Индоевропейская семья включает в себя такие лингвистические группы, как индо-иранцы (Южная Азия), славяне и балты (Восточная Европа), кельты (Западная Европа), германцы (Центральная, Северная Европа).Возможно, у них были и общие генетические предки, которые около 7 тыс. лет назад вследствие миграций оказались в разных частях Евразии, часть ушла на юг и восток (R1a-Z93), положив начало индо-иранским народам и языкам (во многом приняв участие в этногенезе тюркских народов), а часть осталась на территории Европы и положила начало образованию многих европейских народов (R1b-L51), включая славян и русских в частности (R1a-Z283, R1b-L51). На разных этапах формирования уже в древности были пересечения миграционных потоков, что послужило причиной наличия у всех европейских этносов большого количества гаплогрупп. Славянские языки выделились из некогда единой группы балто-славянских языков (предположительно археологическая культура поздней шнуровой керамики). По расчетам лингвиста Старостина это произошло примерно 3,3 тысячелетия назад. Период от V века до н.э. по IV-V век н.э. можно считать условно праславянским, т.к. балты и славяне уже разделились, но самих славян еще не было, они появятся несколько позже, в 4-6 веках н.э. На начальном этапе формирования славян вероятно около 80% составляли гаплогруппы R1a-Z280 и I2a-M423. На начальном этапе формирования балтов вероятно около 80% составляли гаплогруппы N1c-L1025 и R1a-Z92. Влияние и пересечение миграций балтов и славян было с самого начала, потому во многом это деление условно, и в целом отражает только основной тренд, без подробностей. Иранские языки относятся к индоевропейским, и датировка их следующая - древнейшие, от 2-го тысячелетия до н.э. по IV век до н.э., средний - от IV века до н.э. по IX век н.э., и новый - с IX века н.э. по настоящее время. То есть древнейшие иранские языки появляются уже после ухода части племен говоривших на индо-европейских языках из Средней Азии в Индию и Иран. Основными гаплогруппами у них вероятно были R1a-Z93, J2a, G2a3. Западно-иранская группа языков появилась уже позже, примерно в V веке до н.э. Таким образом индо-арии, кельты, германцы и славяне в академической науке стали индоевропейцами, этот термин наиболее адекватный для такой такой обширной и разнообразной группы. Это совершенно корректно. В генетическом аспекте бросается в глаза неоднородность индо-европейцев как по Y-гаплогруппам, так и по аутосомам. Для индо-иранцев характерно в большей степени переднеазиатское генетическое влияние БМАК. Согласно индийским ведам, именно индо- арии пришли в Индию (в Южную Азию) с севера (из Средней Азии), и это их гимны и сказания легли в основу индийских вед. И, продолжая дальше коснемся лингвистики, ведь это русский язык (и родственные ему балтийские языки, например, литовский в составе когда-то существовавшей балто-славянской языковой общности) относительно близок к санскриту наравне с кельтскими, германскими и другими языками большой индо-европейской семьи. Но в генетическом плане индо-арии были уже в большей степени переднеазиатами, по мере приближения к Индии усиливалось и веддоидное влияние. Итак, стало ясно, что гаплогруппа R1a в ДНК-генеалогии - это общая гаплогруппа для части славян, части тюрков и части индо-ариев (т.к. естественно в их среде были представители и других гаплогрупп), часть гаплогруппы R1a1 при миграциях по Русской равнине вошли в состав финно-угорских народов, например мордвы (эрзя и мокша). Часть племен (для гаплогруппы R1a1 это субклад Z93) при миграциях принесли этот индо-европейский язык в Индию и Иран примерно 3500 лет назад, то есть в середине II-го тыс. до нашей эры. В Индии он трудами великого Панини был произведен в санскрит в середине I-го тыс. до н.э., а в Персии-Иране арийские языки стали основой группы иранских языков, древнейшие из которых датируются II-м тысячелетием до н.э. Эти данные подтверждаются: ДНК-генеалогия и лингвистика здесь коррелируют между собой. Обширная часть гаплогруппы R1a1-Z93 еще в древности влились в тюркские этносы и маркируют сегодня во многом миграции тюрков, что и неудивительно ввиду древности гаплогруппы R1a1 , в то время как представители гаплогруппы R1a1-Z280 входили в финно-угорские племена, но при поселении славянских колонистов многие из них были ассимилированы славянами, однако даже сейчас у многих народов например у эрзя по-прежнему доминирующей гаплогруппой является R1a1 -Z280. Все эти новые данные смогла нам предоставить ДНК-генеалогия , в частности, примерные даты миграций носителей гаплогрупп на территории современных Русской Равнины и Средней Азии в доисторическое время. Так ученые всем славянам, кельтам, германцам и т.д. дали имя индоевропейцев, что соответствует действительности с точки зрения лингвистики. Откуда появились эти индоевропейцы? На самом деле индо-европейские языки были задолго до миграций в Индию и Иран, по всей Русской равнине и до Балкан на юге, а до Пиреней на западе. В дальнейшем язык был распространен в Южную Азию - и в Иран и в Индию. Но в генетическом плане корреляций намного меньше. «Единственно оправданным и принятым в настоящее время в науке является применение термина «арии» лишь по отношению к племенам и народам, говорившим на индо-иранских языках». Так в какую сторону шел индо-европейский поток - на запад, в Европу, или наоборот, на восток? По некоторым оценкам индоевропейской языковой семье около 8500 лет. Прародина индо-европейцев до сих пор не определена, но по одной из версий это могло быть Причерноморье- южное или северное. В Индию, как мы уже знаем индо-арийский язык был привнесен около 3500 лет назад предположительно с территории Средней Азии, а сами арии были группой с различными генетическими Y-линиями, как R1a1-L657, G2a, J2a, J2b, H и др. Гаплогруппа R1a1 в Западной и Южной ЕвропеАнализ 67-маркерных гаплотипов гаплогруппы R1a1 из всех стран Европы позволил определить примерный путь миграции предков R1a1 в направлении Западной Европы. И расчеты показали, что почти по всей Европе, от Исландии на севере до Греции на юге, общий предок у гаплогруппы R1a1 был один приблизительно 7000 лет назад!Иначе говоря, потомки как эстафету передавали свои гаплотипы своим же потомкам из поколения в поколение, расходясь в процессе миграций из одного и того же исторического места - которым оказались предположительно Приуралье или Причерноморская низменность. На современной карте - это страны в основном восточной и центральной Европы - Польша, Беларусь, Украина, Россия. Но ареал более древних гаплотипов гаплогруппы R1a1 уводит на восток - в Сибирь. И время жизни первопредка, на которое указывают самые древние, самые мутированные гаплотипы - это 7,5 тысяч лет назад. В те времена еще не было ни славян, ни германцев, ни кельтов. Центральная и Восточная ЕвропаПольша, общий предок R1a1 жил около 5000 лет назад (в основном это субклад R1a1-M458 и Z280). У русско-украинских - 4500 лет назад, что практически совпадает в пределах точности расчетов.Да и даже если четыре поколения - это не разница для таких сроков. В современной Польше гаплогруппы R1a1 в среднем 56%, и в некоторых районах до 62%. Остальные имеют в основном западноевропейскую гаплогруппу R1b (12%), скандинавскую гаплогруппу I1 (17%) и балтийскую гаплогруппу N1c1 (8%). В Чехии и Словакии общий праславянский предок жил 4200 лет назад. Всего ненамного меньше, чем у русских и украинцев. То есть речь идет о расселении на территориях современных Польши, Чехии, Словакии, Украины, Белоруссии, России - все в пределах буквально нескольких поколений, но четыре с лишним тысячи лет назад. В археологии такая точность датировок совершенно немыслима. В Чехии и Словакии потомков гаплогруппы R1a1 около 40%. У остальных в основном западноевропейская R1b (22-28%), скандинавская I1 и балканская гаплогруппа I2a (в совокупности 18%) На территории современной Венгрии общий предок R1a1 жил 5000 лет назад. Там сейчас до четверти потомков гаплогруппы R1a1. Остальные имеют в основном западноевропейскую гаплогруппу R1b (20%) и совокупную скандинавскую I1 и балканскую I2 (суммарно 26%) гаплогруппы. Учитывая, что венгры говорят на языке финно-угорской группы языков, наиболее распространенной гаплогруппой у которых является N1c1 в древних венгерских богатых захоронениях мадьяр в основном находят останки мужчин с гаплогруппой N1c1 , которые и были первыми вождями племен, участвовавших в становлении империи. В Литве и Латвии общий предок реконструируется на глубину 4800 лет. В основном там сегодня субклад Z92, Z280 и M458. Наиболее распространена среди литовцев балтийская гаплогруппа N1c1, достигая 47%. Вообще же для Литвы и Латвии характерен Южно-Балтийский субклад L1025 гаплогруппы N1c1. В общем, ситуация ясна. Добавлю только, что по европейским странам - Исландии, Нидерландов, Дании, Швейцарии, Бельгии, Литвы, Франции, Италии, Румынии, Албании, Монтенегро, Словении, Хорватии, Испании, Греции, Болгарии, Молдавии - общий предок жил 5000-5500 лет назад, точнее установить невозможно. Это - общий предок гаплогруппы R1a по всем перечисленным странам. Общеевропейский предок, так сказать, не считая показанного выше балканского региона, возможной прародины, индоевропейцев около 7500 лет назад. Доля носителей гаплогруппы R1a1 в следующих странах варьируется, от 4% в Голландии и Италии, 9% в Албании, 8-11% в Греции (до 14% в Салониках), 12-15% в Болгарии и Герцоговине, 14-17% в Дании и Сербии, 15-25% в Боснии и Македонии, 3% в Швейцарии, 20% в Румынии и Венгрии, 23% в Исландии, 22-39% в Молдавии, 29-34% в Хорватии, 30-37% в Словении (16% в целом по Балканам), ну и заодно - 32-37% в Эстонии, 34-38% в Литве, 41% в Латвии, 40% в Белоруссии, 45-54% на Украине. В России восточно-европейской гаплогруппы R1a , как я уже упоминал, в среднем 47%, за счет высокой доли балтийской гаплогруппы N1c1 на севере и северо-западе России, но на юге и в центре России доля разных субкладов гаплогруппы R1a достигает 55%. Тюрки и гаплогруппа R1a1Гаплотипы предков везде разные, для разных регионов характерны свои субклады. У народов Алтая и других тюрков также высокие проценты гаплогруппы R1a1, у башкир достигает 40% субклад Z2123. Это дочерняя линия от Z93 и её можно назвать типично тюркской и не имеющей отношения к миграциям индо-иранцев.Сегодня большое количество гаплогруппы R1a1 находиться в Саяно-Алтайском регионе, среди тюркского населения Средней Азии. У киргизов достигая 63%. Ни русскими ни иранцами их не назовешь. Получается назвать всю гаплогруппу R1a1 единым названием - грубое утрирование, по меньшей мере, а по большей - невежество. Гаплогруппы - это не этносы, на них не записана лингвистическая и этническая принадлежность носителя. К генам тоже прямого отношения гаплогруппы не имеют. Для тюрков в основном характерны различные субклады Z93, но в Поволжье встречаются и R1a1-Z280, возможно перешедшие к поволжским тюркам от поволжских финнов. Гаплогруппа R1a1-Z93 также в умеренной частоте характерна для арабов, для левитов - подгруппы ашкеназских евреев (у последних был подтвержден субклад CTS6). Данная линия уже на самых ранних этапах приняла участие в этногенезе этих народов. Территория первоначального распространения гаплогруппы R1a1 в Европе - это, вероятно, территория Восточной Европы и возможно Причерноморская низменность. До этого вероятно в Азии, возможно в Южной Азии или Северном Китае. Кавказские гаплотипы R1a1Армения. Возраст общего предка гаплогруппы R1a1 - 6500 лет назад. В основном тоже субклад R1a1-Z93, хотя есть и R1a1-Z282.Малая Азия, Анатолийский полуостров. Исторический перекресток на путях между Ближним Востоком, Европой и Азией. Это был первый или второй кандидат для «индоевропейской прародины». Однако общий предок гаплогруппы R1a1 жил там около 6500 лет назад. Ясно, что этой прародины, судя по гаплотипам, в Анатолии быть практически может, либо первоначальными индо-европейцами были носители гаплогруппы R1b . Но высока вероятность низкой представленности в общей базе гаплотипов индивидов из Турции. Итак, и армяне, и анатолийцы - у всех предок или тот же самый, или предки очень близки по времени, в пределах нескольких поколений - это субклад Z93 и Z282*. Следует отметить, что 4500 лет до общего предка гаплогруппы R1a1-Z93 в Анатолии хорошо согласуется со временем появления хеттов в Малой Азии в последней четверти III тысячелетия до н.э, хотя многие линии R1a1-Z93 там могли появиться после миграций тюркских народов на полуостров уже в нашей эре. Алексей Зоррин *** . | |
В данном исследовании используется более 80 000 аутосомных снипов, реперных точек - это очень высокое разрешение, позволяющее уловить даже относительно небольшие влияния на генетическом уровне у основной массы народа. Данные сравнительного анализа взяты из открытого исследования В.Веренича - специалиста по сравнительному анализу генетических компонент. Сами генетические калькуляторы находятся на сервисе GedMatch , и позволяют любому желающему узнать свое сравнительное положение на генетическом графике. Для этого достаточно иметь результаты аутосомного теста компании FTDNA , либо 23andMe. В конце исследования приведены карты географического распространения и частотных максимумов для основных аутосомных компонент из проекта MDLP World-22 .
На графиках приведенных ниже показаны основные компоненты и их среднее процентное соотношение для каждой из популяций. В одной строке - процентная разбивка для одной популяции. Каждое деление (вертикальная черта) соответствует 10%, а названия аутосомных компонент располагаются в той же последовательности слева направо, что и в легенде сверху вниз. Чем более похож процентный состав общей генетики у разных народов, тем более похожим выглядит рисунок на приведенном графике. Итак приступим...
Генетика немцев, литовцев, русских, шведов, финнов и др.
На данном графике приведены основные генетические компоненты для европейских народов и выравнены по уменьшению восточно-европейской компоненты (North-East-European) в различных популяциях. Как видно все европейские народы достаточно различны в генетическом плане, и имея в своем наборе одинакового происхождения генетические компоненты, они тем не менее в весьма различных процентах. Для всех славян и балтов в целом одной из максимально-значимых является данная компонента Восточной Европы, которая в максимуме у литовцев и белорусов. Вероятно со времен археологической "культуры шнуровой керамики" территория этих стран была центром происхождения данной компоненты. Она представлена более 80% у литовцев, и всего 20% у итальянцев.Фиолетовый цвет обозначает атланто-средиземноморскую компоненту, и она увеличивается по мере движения с севера-востока на юго-запад. Так у финнов она достигает в среднем 15%, а у итальянцев 40%. Остальные компоненты менее выражены.
Генетика русских украинцев белорусов

На данном графике приведены восточные славяне - русские, белорусы, украинцы . Обращает на себя внимание сходство генетических рисунков трех перечисленных народов, и в пределах погрешности они различаются совсем незначительно - у украинцев и южных русских небольшое повышение западно-азиатской компоненты, а у северных русских - небольшой рост одной из Сибирских компонент, названной условно самодийской, и повышение компоненты мезолита Европы примерно до 10%, что по последнему показателю сближает их скорее с германоязычной популяцией Скандинавии - шведами.
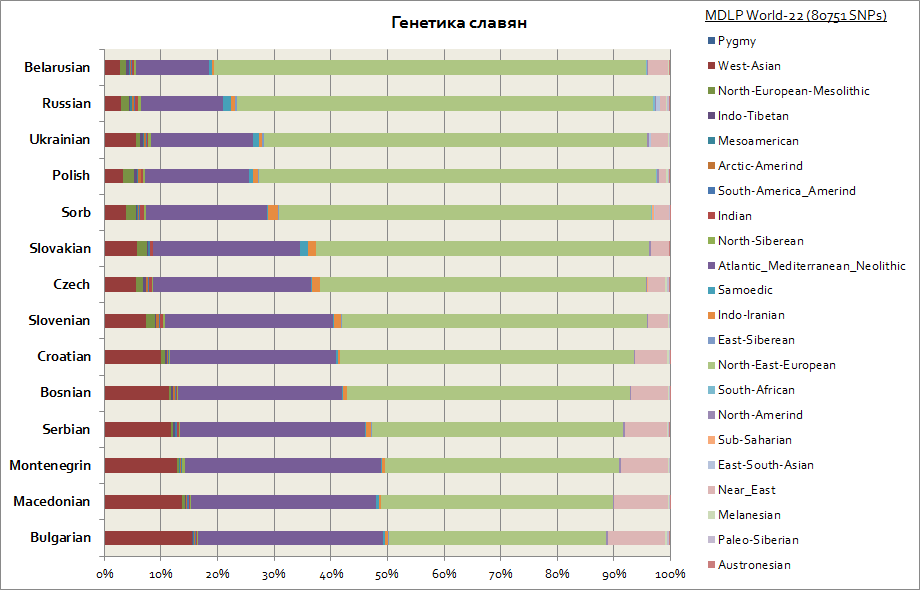
На этом графике изображены все славяне, включая западных - поляков и чехов, а также южных - сербов, болгар, македонцев и т.д.
Основных компонент у всех славян - 2. Это восточно-европейская и атланто-средиземноморская. Первая в максимуме у белорусов, а вторая у всех южных славян - сербов,
македонцев, болгар. Восточно-европейская компонента более первична по происхождению у славян, а атланто-средиземноморская является в большей степени приобретенной в процессе
миграций славян на Балканы. У западных украинцев и словаков слабое повышение Самодийской компоненты относительно соседних славянских народов - белорусов, чехов, поляков; вероятно это генетических след средневековых миграций гуннов и угров в Центральную Европу.
Генетика славян, русских и татар, германцев, кавказцев, евреев и др.
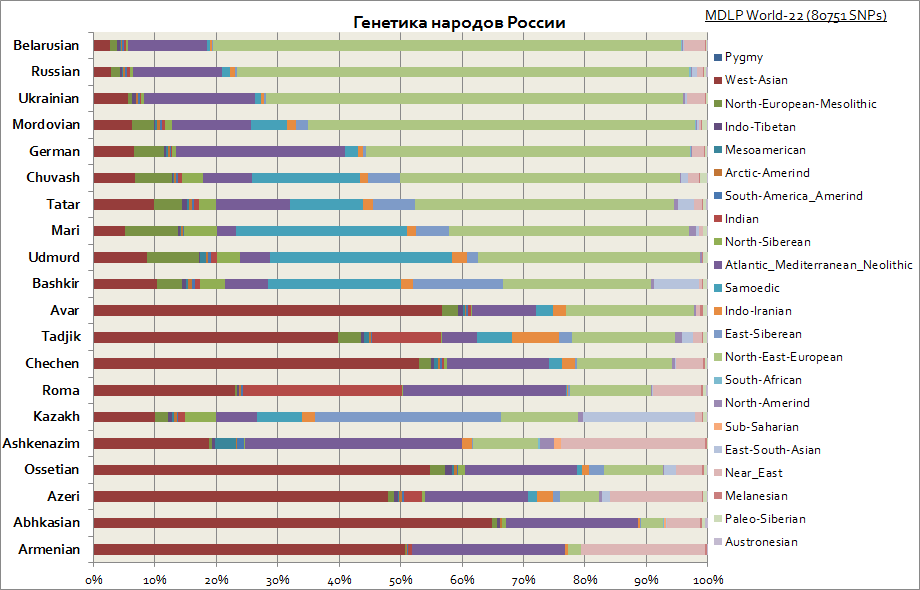
Данный график отображает различное происхождение среди народов России. Как видно у славян основной является восточно-европейская компонента, а у народов Поволжья повышается доля сибирских компонент. В то время как для кавказцев наиболее характерны западно-азиатская компонента, средиземноморская и ближне-восточная.
Генетика финнов, угров, удмуртов, венгров, саамов и др.

Как видно для финнов, вепсов и карел характерно схожее генетическое происхождение со славянами. У них также наиболее максимальна восточно-европейская компонента, уменьшается ближе к Уралу и Поволжью, с увеличением в этом регионе сибирских компонент. Также у всех финно-угорских народов выражена значительно компонента мезолита Европы, которая достигает почти 80% у саамов и связана с до-индоевропейским и до-неолитическим населением Европы. Для венгров в целом характерен набор тех генетических компонент, что и для других популяций карпатского региона и Центральной Европы.
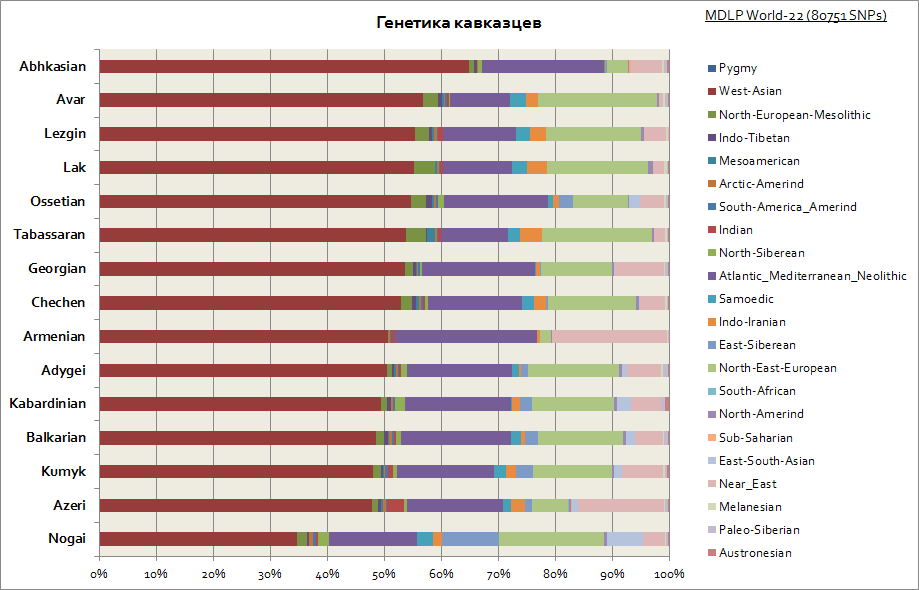
Как видно для всего Кавказа характерно сравнительно схожее генетическое происхождение - это большая доля западно-азиатской компоненты и средиземноморской. Немного выбиваются только ногайцы - у них повышается доля сибирских компонент.

Как видно у ашкеназов и у сефардов высокая частота западно-азиатской, атланто-средиземноморской и ближневосточной компонент. При этом у ашкеназов незначительное повышение сибирской компоненты, что вероятно связано с хазарским наследием и рост до 30% Восточно-Европейской компоненты, что по данному показателю сближает их со странами юга Европы.
Особенно выбиваются из их "компании" только эфиопские евреи и индийские евреи. У первых высока доля субсахарской Африки (до 40%), а у вторых доля южно-азиатской генетической компоненты, условно названной Индийская (до 50%).
Генетика татар, башкир, азербайджанцев, чувашей и др.
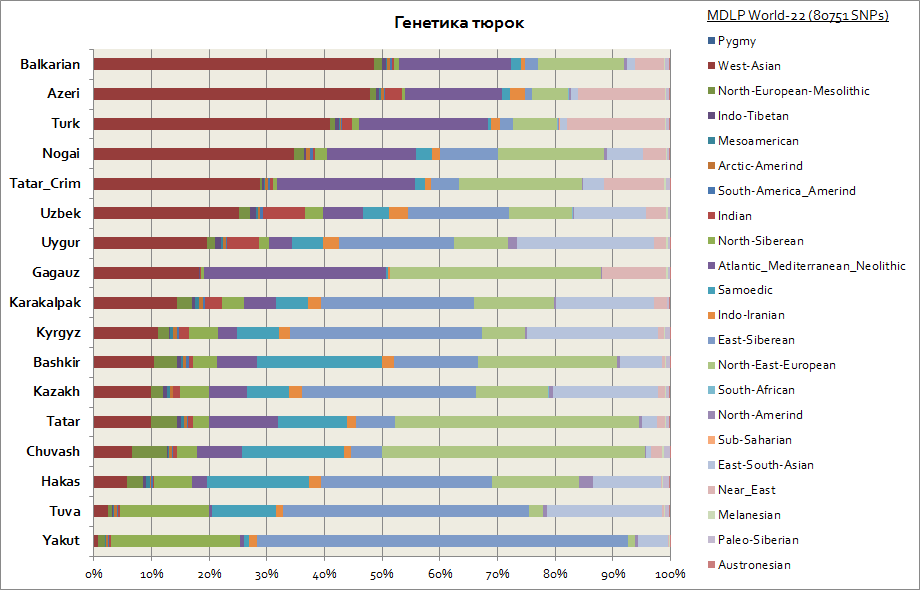
Тюрки в генетическом плане оказались одним из наиболее неоднородных этносов, потому генетические компоненты у них значительно различаются. Так, учитывая, что первичная родина тюрков - это Сибирь, то такие народы как якуты, тувинцы, хакасы - сохранили в наибольшем проценте Восточно-сибирскую аутосомную компоненту, которая достигает у них от 30 до 65%. Данная генетическая компонента также является основной у киргизов и казахов. Остальные компоненты сближают тюрков с народами из регионов проживания. Так, для якутов и тувинцев - это Северо-сибирская и Самодийская компоненты. В сумме данные 3 сибирские компоненты у якутов составляют до 90%, у тувинцев до 70%, с возрастанием до 20% восточно-юго-азиатской компоненты, которая связана в большей степени с миграционными потоками населения Восточной Азии. Для башкиров доля 3-х сибирских компонент составляет до 45%, и юго-восточно азиатской до 10%. У татар данные 3-х сибирских генетических компонент в среднем от 25 до 50%. При этом доля компонент, характерных для европеоидного населения у башкир составляет до 45%, а у татар в среднем от 50 до 70%. Генетика азербайджанцев и турков в пределах погрешности практически не различается, у них как и у остальных народов региона Кавказа и Закавказья значительно представлены Западно-Азиатская компонента (достигает 50%) и атланто-средиземноморская (в среднем до 20%). Доля 3-х сибирских компонент представлена у азербайджанцев, турков и балкарцев - на уровне 3-7%.
Заключение
Генетика народов не имеет прямой и значительной корреляции с распространением языковых семей, или с процентом однородительских маркеров - гаплогрупп Y-ДНК и мт-ДНК , представленных у конкретной популяции. Наибольшая корреляция прослеживается по территориально-географическому принципу. Так, доля Сибирских компонент, характерных в целом для монголоидной расы плавно уменьшается с Востока на Запад, а доля компонент, характерных для европеоидной расы - соответственно увеличивается. В пограничных территориях по линии от севера Урала до Центральной Азии примерно равное их соотношение. В регионах восточнее Байкала уже практически не представлены генетические компоненты, характерные для большой европеоидной расы, в тоже время в регионах, западнее линии Печора-Поволжье сходят на нет сибирские компоненты, характерные для большой монголоидной расы.Распространение Восточно-европейской генетической составляющей в Сибирь, произошло по большей степени уже в бронзовом веке (культуры Андроновского круга), хотя отдельные пики на крайнем востоке Сибири у чукчей, возможно, связаны уже с миграциями русских в XVII веке.
Доля Субсахарской компоненты, характерной для негроидной расы распространена по всей Африке - вплоть до южного Средиземноморья и Северной границы Африканского континента, достигая максимума в экваториальной его части, а за его пределами уже практически не встречается; легким фоном распространена на Аравийском полуострове и южной части Иранского плато.
География генетических компонент
Алексей Зоррин
Проект
Строго говоря, никакого славянского гена, точно так же как тюркского или финского, или германского или какого угодно другого гена, не существует. Гены -- структурно-функциональные единицы генетического материала, наследственный фактор, который можно условно представить как отрезок молекулы ДНК – намного древнее, чем любой народ на земле. Тем не менее, генетики выделяют гаплогруппу, которая характерна для представителей славянских народов. Гаплогруппой называют набор нуклеотидов мужской Y-хромосомы, который передается из поколения в поколение без каких-либо изменений в течение тысячелетий. Передается она исключительно по мужской линии. Так вот, приблизительно четыре с половиной тысячи лет назад на Среднерусской равнине родился мальчик с несколько иной, чем у отца гаплогруппой. Генетическая классификация гаплогруппы отца выглядит так: R1a. Мутировавшей гаплогруппе сына современные генетики присвоиликлассификацию R1a1. Эта мутация оказалась очень живучей. И в настоящее время обладатели гаплогруппы R1a1 составляют 70% населения России, Белоруссии и Украины, а так же являются большинством в других славянских странах. Собственно, этот самый набор нуклеотидов и есть, в известном смысле, биологический маркер славянства. Но, разумеется, в наши дни не может быть и речи о неких «чистых» народах, избежавших смешения с другими этносами. Поэтому и славяне тоже весьма неоднородны. Наряду с этой самой гаплогруппой R1a1, свой генетический след в них оставили многие народы. Среди русского населения, действительно, около 14% является носителями гаплогруппы, характерной для финно-угров, что совершенно естественно, ведь финно-угорские племена – древние обитатели земель, где сейчас располагается Россия. Зато гаплогруппа, характерная для монголов, вопреки расхожей поговорке «поскреби русского и найдешь татарина», у русских встречается крайне редко – всего от полутора до трех процентов, у украинцев тоже немного – около пяти процентов. Зато среди украинцев почти 37% населения являются обладателями гаплогрупп, характерных для Балкан, что, опять же, вполне естественно в силу географической близости и частых контактов. Есть свои особенности и у жителей других славянских стран. В Белоруссии, например, есть немало носителей гаплогрупп, характерных для народов балтийской группы, чехи и другие западные славяне ближе к западноевропейским народам, у болгар имеется изрядный фракийский след, которым одарили пришлое южнославянское племя древнейшие жители этого региона. Народ определяют не гены, а язык, традиции, религия и все остальное, что мы называем культурой. Поэтому понятие «славянский ген» следует относить все же к области поэтических метафор, а не науки
Реконструкция генетической и лингвистической истории балто-славянских популяций
Эта наиболее полная работа по генофонду славянских и балтских народов подводит итоги многолетних исследований. Для реконструкции давней истории народов, говорящих на родственных языках, применен междисциплинарный подход. Генетики и лингвисты проследили пути формирования генофонда всех групп славян и балтов одновременно по трем генетическим системам: по Y-хромосоме (отцовские линии наследования), по митохондриальной ДНК (материнские линии наследования) и по широгеномным данным об аутосомных маркерах (где отцовские и материнские линии представлены равноправно). Прослежено, какие местные популяции впитывал генофонд славян при их расселении по Европе: именно этот глубинный субстрат сформировал основные различия генофондов разных ветвей славян. Корреляция генетического разнообразия с лингвистическим оказалась велика, но еще больше — с географическим соседством популяций. Результатом исследования стало и уточнение древа балто-славянских языков.
Отклики на исследование в СМИ и на научно-популярных сайтах — в конце текста
Формирование генофонда балто-славянских популяций исследовала большая международная группа генетиков и лингвистов. Статья с результатами их работы опубликована в журнале PLoS ONE . Исследование проведено под руководством доктора биол. наук О.П. Балановского (Институт общей генетики и Медико-генетический научный центр) и академика Рихарда Виллемса (Эстонский биоцентр и Тартусский университет). В нем участвовали исследователи из многих стран, в которых славянские и балтские народы составляют большинство населения — России, Украины, Белоруссии, Литвы, Хорватии, Боснии и Герцеговины, а также ученые Эстонии, Великобритании и консорциум международного проекта Genographic. Это наиболее полная работа по генофонду славянских и балтских народов подводит итог многолетним исследованиям многочисленных авторов статьи и учитывает данные других научных коллективов.
На балто-славянских языках говорит примерно треть современного населения Европы, а по площади балтские и славянские народы занимают около половины Европы. Лингвисты сходятся во мнении, что балтские и славянские языки не только родственны, но и имеют общий корень в семье индоевропейских языков. По их оценкам прото-балто-славянский язык отделился от других индоевропейских языков в интервале от 7000 до 4500 лет назад, и произошло это, вероятнее всего, в Центральной Европе. Расхождение балтской и славянской языковых ветвей датируется временем 3500-2500 назад. Дальнейшее разделение славянских языков происходило уже относительно недавно — 1700-1300 лет назад. С ранним средневековьем (примерно 1400-1000 лет назад) связывается так называемая «славянизация Европы» — период быстрого распространения славянских языков на огромных территориях. В Восточной Европе славяне распространялись на территории, где проживали балтские, финно-угорские и тюркские популяции, в Западной Европе – на территории носителей германских языков, на Балканах – на территории местных разноязыких популяций.
Но как эти изменения в культуре Европы, фиксируемые распространением славянских языков, повлияли на генофонд Европы? Именно это стало главным вопросом исследования. Ведь до сих пор оставалась недостаточно изученной генетическая история балто-славянских популяций и их взаимодействие с генофондами популяций, говоривших на других языках — финно-угорских, германских, тюркских.

Что изучали
Для максимально полного изучения балто-славянских популяций ученые использовали все три генетические системы, которые на данный момент наиболее информативны для исследования генофонда.
1) Y-хромосома, которая наследуется по отцовской линии: изучено 6078 образцов из 62 популяций;
2) Митохондриальная ДНК (мтДНК), которая наследуется по материнской линии: изучено 6876 образцов мтДНК из 48 популяций;
3) Широкогеномные (полногеномные) маркеры: 1 297 образцов из 16 популяций. Это точки генетического разнообразия (однонуклеотидного полиморфизма, SNP), которые разбросаны по всему геному, и расположены на аутосомах – неполовых хромосомах.
Для анализа ADMIXTURE использовано 200 тысяч SNP-маркеров, которые являются общими для трех использованных панелей Illumina (610K, 650K и 660K) и несцепленными друг с другом; для анализа общих фрагментов были использованы все 500 тысяч маркеров, которые являются общими для трех использованных панелей Illumina (включая маркеры, сцепленные друг с другом); для анализа главных компонент и расчета генетических расстояний использованы 57 тысяч маркеров, которые являются общими для панелей Illumina и Affimetrix, и при этом не сцепленными друг с другом.
Значительная часть этих внушительных массивов данных получена авторами впервые — 1254 образцов по Y-хромосоме, 917 образцов по мтДНК, 70 образцов по широкогеномным маркерам. Остальные данные взяты из ранее опубликованных работ. Для сравнения использованы все накопленные к нынешнему времени данные по другим генофондам Европы.
По всем трем генетическим системам были изучены практически все современные народы, говорящие на языках балто-славянской группы – шестнадцать народов по единой обширной панели маркеров:
балтские народы — латыши и литовцы;
восточные славяне — белорусы, русские, украинцы;
западные славяне — кашубы, поляки, словаки, сорбы, чехи;
южные славяне — болгары, боснийцы, македонцы, сербы, словенцы, хорваты.
Такие подробные и разносторонние данные по какой-либо группе народов (охват всех этносов, да еще и по всем основным генетическим системам) являются большой редкостью в популяционных исследованиях. Поэтому они позволяют решить не только конкретную, но и более общую методологическую задачу. Конкретная задача – это описание генофонда самих славян и балтов, а общая – на их примере изучить, как связаны друг с другом разные признаки, по которым обычно характеризуются популяции: Y-хромосомное разнообразие, митохондриальное разнообразие, полногеномное разнообразие, лингвистическое родство, географическое положение популяций.
Генетический ландшафт славян через три призмы

Генетические соотношения друг с другом всех изученных популяций, установленные в результате исследования, показаны на рисунках.
Рис. А представляет результаты для широкогеномных (аутосомных) SNP-маркеров. Эти маркеры называются аутосомными потому, что находятся на неполовых хромосомах (аутосомах). А широкогеномными они называются потому, что равномерно разбросаны по всему геному.
Рис. В представляет результаты по Y-хромосоме, полученные на основании частот ее гаплогрупп.
Рис. С отражает результаты, полученные по частотам гаплогрупп митохондриальной ДНК (мтДНК).
Чтобы показать относительную близость и удаленность разных популяций на двумерном графике, в популяционной генетике используются два метода, заимствованные из многомерной статистики: метод главных компонент и метод многомерного шкалирования. По своей сути они близки, но достоинства и недостатки их противоположны. Метод главных компонент показывает положение популяций математически точно, но теряет порой значительную часть генетической информации, содержащейся в исходных данных. А метод многомерного шкалирования, наоборот, использует всю генетическую информацию, но геометрические расстояния между популяциями-точками на графике могут быть несколько искажены относительно рассчитанных генетических расстояний между ними. В данном случае для аутосомных данных был применен метод главных компонент, а для Y-хромосомных и митохондриальных данных – метод генетических расстояний.
Как видно, и по широкогеномным маркерам, и по Y-хромосоме (А и В) большинство балто-славянских популяций выстраиваются вдоль оси север-юг.
Восточные славяне – русские, белорусы и украинцы — отчетливо группируются, Они образуют свой кластер, хотя внутри него русские, белорусы и украинцы не перекрываются полностью друг с другом. Исключение составляют северные русские, которые генетически отдалены от остальных восточных славян и тяготеют к соседним финно-угорским популяциям.
Из западных славян чехи и в меньшей степени словаки отличаются от восточных славян и смещены в сторону немцев и других западноевропейских популяций. А вот поляки наиболее близки к восточным славянам. Фактически на графиках поляки, русские, белорусы и украинцы формируют общий кластер, а словаки и особенно чехи несколько удалены от него.
Южные славяне формируют на графике дисперсную группу, которая внутренне поделена на западный (словенцы, хорваты и боснийцы) и восточный (македонцы и болгары) регионы с сербами посередине. При этом словенцы генетически близки к венграм (географически близкий, но не славянский народ), а восточная ветвь южных славян тоже группируется с неславянскими, но географически близкими румынами и до некоторой степени с греками.
Балтские народы — латыши и литовцы — обнаруживают генетическую близость к эстонцам, говорящим на языке финно-угорской группы, и к некоторым восточным славянам (белорусам). Оказалось также, что балтские популяции близки к волжской группе финно-угорских народов (особенно к мордве). Авторы уточняют, что это может отражать исторические события – в древности ареал балто-язычных популяций простирался далеко на восток и почти доходил до нынешнего ареала мордвы.
Важно, что все перечисленные закономерности выявлены по независимым и, казалось бы, совершенно разным генетическим системам – Y-хромосоме и широкогеномным аутосомным маркерам.
По мтДНК (рисунок С), как обычно, степень структурированности генофонда выражена значительно слабее, что связано с более низким филогенетическим разрешением в имеющихся данных по мтДНК. Но, хотя и не так четко выраженные, в результатах по мтДНК проглядывают те же самые закономерности. Например, и на графике по мтДНК большинство восточно-славянских популяций перекрываются друг с другом, северные русские отделены от них, а южные славяне генетически сходны со своими неславяноязычными соседями по Балканам.
Сравнивая степень выраженности одних и тех же закономерностей в результатах по разным генетическим системам, авторы подчеркивают, что Y-хромосома часто выявляет закономерности более детально, чем не только мтДНК, но и более модные широкогеномные маркеры.
Поиск глубинных предков
Чтобы сравнить популяции по составу их предковых компонентов, часто используется программа ADMIXTURE («смешение», или «состав»). В нее закладывают широкогеномные данные по большому числу популяций и задают число гипотетических предковых популяций, из которых сформировались все эти современные популяции. Программа вычисляет, каков должен быть генетический состав этих предковых популяций (предковых компонентов), и рисует для каждой современной популяции цветной спектр, указывающий на доли этих предков в ее генофонде. Понятно, что такая модель достаточно условна — в реальности вряд ли современные генофонды сформировались в результате смешения фиксированного заданного числа предковых популяций. Но такая упрощающая модель часто оказывается полезна, а выявляемые предковые компоненты обычно имеют реальный смысл. Например, при анализе данных в масштабе мира всегда первым выделяется африканский компонент, который составляет почти 100% в африканских популяциях южнее Сахары, а его доля в других популяциях мира хорошо соответствует степени их прямого или опосредованного смешения с популяциями Африки.
В данной работе также был применен метод ADMIXTURE – авторы задавали разное число предковых популяций и опубликовали все соответствующие графики, но специальный тест показал, что статистически наиболее обоснованные результаты получены в случае, когда число предковых компонентов задавалось равным шести (К=6). В этом случае авторы получили вот такую картину.

У балто-славянских популяций почти весь спектр представлен двумя цветами: синим (предковый компонент k3) и голубым (предковый компонент k2), хотя и в разных пропорциях. Если посмотреть на Европу в целом, то видно, что k3 (синий) вносит большой вклад во все европейские популяции и снижается от северо-востока к югу. Этот предковый компонент максимален у балтских популяций, превалирует у восточных славян (80-95%) и снижается у южных славян (55-70%). Напротив, k2 (голубой) более характерен для популяций средиземноморского и кавказского регионов и снижается к северу Европы. У южных славян на него приходится примерно 30%, у западных славян он снижается до 20%, а у северных русских и балтских популяций — до 5%.
Видно, что у славян есть еще лимонно-желтый цвет в предковом спектре, это компонент k5, который представлен сколько-нибудь значимо только у восточных славян, а из них больше выражен у северных русских. По происхождению этот компонент сибирский, поскольку, как видно на графике, он составляет основную часть спектра для популяций Сибири. А вот компонент k6 (темно-желтый), который доминирует в Китае, Монголии и на Алтае, у русских почти на нуле. Это означает, что восточный след в генофонде северных русских связан, скорее, с древними миграциями из лесов и тундр Сибири, чем из степей Центральной Азии (новое опровержение расхожего представления о большом влиянии на русский генофонд татаро-монгольского ига). Темно-зеленый компонент k4 характеризует популяции Южной Азии, распространен также на Ближнем Востоке и Средиземноморье. Поэтому неудивительно, что он, пусть с небольшой частотой, но встречается у южных славян и других народов Балканского полуострова, но почти сходит на нет у западных и восточных славян.
Из рассмотрения состава предковых компонентов следует вывод о значительном генетическом сходстве большинства западных и восточных славян на большой территории — от Польши на западе до европейской части России на востоке. А южные славяне, географически ограниченные небольшим Балканским полуостровом, существенно отличаются от западных и восточных.
Но как возникли эти отличия?
Общие фрагменты геномов славян и их соседей
Чтобы ответить на этот вопрос, авторы провели тонкий анализ генофонда для двух групп славян: в первую вошли западные и восточные славяне (ведь они генетически оказались очень схожи), а во вторую – южные славяне. Сравнение проводили по наличию одинаковых фрагментов хромосом у людей, происходящих из этих групп популяций. Этот метод называется IBD анализ – его название происходит от классического понятия популяционной генетики «identical by descent», то есть поиск генетических фрагментов, идентичных по происхождению. Эти фрагменты разными людьми, представителями разных популяций, унаследованы от одного и того же общего для них предка. Понятно, что чуть ли не в любой популяции мира может найтись хоть один потомок представителя западных и восточных славян, и, наоборот, среди восточных славян может найтись хоть один потомок чуть ли не любого народа мира. Но это единичные совпадения – поэтому популяционная генетика и изучает популяции, а не отдельных ее представителей. Те популяции, в которых таких совпадений найдено много, находятся действительно в значительном родстве друг с другом, точнее, имеют значительное число общих предков. Эти общие фрагменты, по сути, являются гаплотипами, похожими на гаплотипы мтДНК и Y-хромосомы тем, что также имеют одного предка, но отличными тем, что с ходом времени разбиваются рекомбинациями — обменом участками между хромосомами, пришедшими от отца и от матери, при делении клетки. А гаплотипы дают возможность датировок и по аутосомным маркерам – зная скорость рекомбинаций, можно по длине сохранившихся общих гаплотипов оценить, сколько времени прошло от общих предков, то есть давно ли существовала общность генофондов.
Было подсчитано число общих гаплотипов между «западно-восточными» славянами (авторам пришлось пользоваться этим неуклюжим термином за неимением лучшего) и восемью другими группами народов Европы:
1) южными славянами (болгары, боснийцы, македонцы, словенцы, хорваты);
2) популяциями Западной Европы (итальянцы, немцы, французы);
3) балтскими популяциями (латыши, литовцы);
4) популяциями северо-восточной Европы (западно-финские народы – вепсы, карелы, финны, эстонцы);
5) популяциями центральной Европы, ареал которых находится между западно-восточными и южными славянами — их авторы условно называют «между-славянскими популяциями»; это удивительно разноязычные популяции: гагаузы говорят на языке тюркской группы алтайской языковой семьи, венгры – на языке угорской группы уральской языковой семьи, а румыны – языке романской группы);
6) греками;
7) популяциями волжского региона и Приуралья (башкиры, коми, мордва, татары, удмурты, чуваши);
8) северокавказскими популяциями (адыги, балкарцы, ногайцы).
Если принять за эталон число общих гаплотипов между западно-восточными и южными славянами, то часть окружающих неславянских популяций будет (по числу общих гаплотипов) выше этого эталона, часть ниже, а часть равняться ему. Ниже эталона (то есть имеют меньшее родство с западно-восточными славянами, чем южные славяне) оказались народы Поволжья, Западной Европы, Кавказа, а также греки.
Казалось бы, можно говорить о большем родстве славянских генофондов друг с другом, чем с окружающими неславянскими народами. Отчасти это так, но все не так просто – в два раза выше эталона оказалось родство генофондов балтов и популяций северо-восточной Европы (вепсы, карелы, финны, латыши, литовцы, северные русские, эстонцы). Можно впасть в противоположную крайность и считать, что «западно-восточные» славяне генетически родственны не южным славянам, а только своим географическим соседям, вероятно, за счет ассимиляции родственных им народов. Но картину дополнительно осложняет то, что с народами, живущими сейчас на территориях посередине между «западно-восточными» и южными славянами – то есть с венграми, румынами и гагаузами — у западно-восточных славян число общих фрагментов генома такое же, что и с южными славянами (эти «между-славянские» популяции находятся на уровне эталона).
Поэтому авторы провели еще один аналогичный анализ, но теперь поставив в центр рассмотрения южных славян. Сравнивалось число общих генетических фрагментов у них и окружающих групп популяций. Оказалось, что число общих фрагментов у южных славян с «западно-восточными» славянами примерно такое же, что и число их общих фрагментов с «между-славянскими» популяциями (гагаузы, венгры, румыны). А вот число общих фрагментов с географически соседними греками значительно меньше. Учтем, что западно-восточные славяне географически дальше от южных славян, чем «между-славянские», поэтому с точки зрения географии число общих фрагментов с «западно-восточными» славянами должно было бы быть меньше. А раз это не так, значит языковое родство «западно-восточных» и южных славян отчасти проявляется и при этом анализе общих фрагментов генома. Тем более, что, хотя общие фрагменты генома, найденные между двумя группами славян и разнятся по длине, фрагментов длиной около 2-3 сантиморганид чуть больше, чем других, а именно такой длины фрагменты и должны были сохраниться со времени славянской экспансии второй половины I тысячелетия н.э.
Эти результаты по славянам, из которых нельзя сделать однозначных выводов, следует сравнить с недавним похожим исследованием тюркоязычных популяций (Yunusbaev et al., 2015). Казалось бы, в обоих случаях идет быстрое распространение носителей языков (соответственно, тюркских или славянских) по обширным территориям, которое не может не сопровождаться ассимиляцией местного (дотюркского или дославянского) населения. Но в случае с тюрками метод анализа общих фрагментов выявил – пусть очень небольшой — компонент генома, который тюрки принесли со своей вероятной алтайской прародины. А в случае славян картина оказалась гораздо более сложной. Возможно, это связано с тем, что тюрки в ходе расселения часто ассимилировали генетически резко отличные от них и друг от друга популяции, а славяне распространялись по территории Европы с ее относительно гомогенным генофондом, и часть ассимилированных ими популяций была родственна, по крайней мере, балтским группам.
В целом, из этого анализа общих фрагментов можно сделать два вывода. Прежде всего, явно видны результаты смешения генофонда западно-восточных славян с другими популяциями северной части Восточной Европы. Во-вторых – пусть и далеко не столь выразительно – видна и несколько большая степень родства западно-восточных и южных славян друг с другом, чем можно было бы ожидать, исходя просто из географического расстояния между ними.
Схема и результаты анализа общих фрагментов генома (IBD)


Реконструкция дерева языков.
В коллектив авторов входили не только генетики, но и ведущие российские лингвисты. Это позволило использовать в этой работе уточненное древо родства балто-славянских языков благодаря обновлению и перепроверке массива лексикостатистических данных. Лексикостатистика занимается выявлением скорости языковых изменений и определением времени разделения родственных языков и степени родства между ними. Исходным материалом послужили лексические списки (списки Сводеша) 20 современных балто-славянских языков и диалектов.

После разделения балтской и славянской ветвей самая первая развилка на славянской ветви оказалась тройной – разделение славян на западную, восточную и южную ветви – и датированной около 1900 лет назад. Дальнейшее разделение славянских языков началось в V-VI веках (около 1300-1500 лет назад): восточная ветвь разделилась на русский и украинский/белорусский, западная ветвь – на чешско/словацкий, протосорбсколужицкий и польский/кашубский, южная ветвь – на сербско-хорватский, болгарский, македонский. Выделение современных языков произошло 1000-500 лет назад. Такая датировка дерева соответствует историческим и археологическим данным, которые говорят о быстром распространении славян по Европе во второй половине 1-го тысячелетия н.э.
Генетическое разнообразие на разных уровнях лингвистического дерева
Поскольку лингвистическое дерево славянских языков так точно построено, появилась возможность проанализировать, как распределено по этому дереву генетическое разнообразие славянских популяций, оцененное по частотам гаплогрупп Y-хромосомы. Этот анализ проведен с помощью стандартной процедуры теста AMOVA.
Оказалось, что генетические различия между популяциями, говорящими на одном и том же языке, хотя и варьируют от почти нулевых значений (для говорящих на чешском или македонском) до значения 0,05 (для говорящих на северных диалектах русского языка), в среднем составили только 0,01.
Далее, частоты во всех этих популяциях одного народа были усреднены и получены среднеэтнические частоты гаплогрупп. А затем были рассчитаны генетические различия между этими среднеэтническими характеристиками народов в пределах каждой ветви славянских языков. Эти различия оказались не совсем одинаковыми для разных ветвей: например, для западных славян различия больше, чем для восточных, но это и можно было ожидать, глядя на графики их генетических взаимоотношений. Однако в среднем различия между этносами оказались больше – 0,03.
Наконец, были рассчитаны средние частоты гаплогрупп для трех ветвей славянских языков – западных, восточных и южных — и различия между ними возросли еще в два раза — около 0,06.
Согласно принципу эквидистантности, разработанному отечественной школой геногеографии, если система популяций развивается сама по себе, без больших внешних влияний, то постепенное разделение популяций приводит к линейному накоплению и лингвистического, и генетического разнообразия. В результате генетическое разнообразие примерно одинаково на всех уровнях – что между популяциями одного народа, что между этносами одной ветви, что между разными ветвями (их усредненными характеристиками). Действительно, ведь предки разных групп славян некогда были лишь близкими друг к другу популяциями одного народа, и их языки отличались не больше, чем сейчас отличаются диалекты одного языка. А усредняя частоты по всем современным популяциям ветви, мы как бы находим ее центр тяжести, точку происхождения, реконструируем генофонд этой предковой популяции.
Но все это, как сказано выше, работает лишь тогда, когда популяции предоставлены сами себе и мало взаимодействуют с соседями. Однако для славян величины генетического разнообразия на разных иерархических уровнях не одинаковы: при эквидистантности они должны быть 0.01, 0.01, 0.01, а они резко различаются — 0.06, 0.03, 0.01. Это говорит о том, что популяции славян как раз активно смешивались с окружающими народами. А то, что наибольшая изменчивость приходится на самый древний уровень (различия между тремя ветвями славянских языков) указывает, что эти взаимодействия были особенно сильными на ранних этапах истории славянских популяций.
Родство или соседство?
Сравнить роли, которые сыграли география и языки в формировании генетического разнообразия балто-славянских популяций, можно с помощью теста Мантеля. География играет двойную роль. Конечно – это фактор географического соседства, который сближает генофонды через смешанные браки между соседями. Но с другой стороны, география может отражать и происхождение, когда родственные народы не уходят далеко друг от друга, а расселяются на соседние территории. Языки – это фактор изначального родства генофондов или частей генофондов, унаследованных от общих предков вместе с общим языком (или не унаследованных, если язык сменили, а генофонд остался почти прежним).
Тест был независимо проведен для трех генетических систем: Y-хромосома, мтДНК и аутосомные маркеры. Все три варианта теста показали чрезвычайно высокую корреляцию между генетикой и географическим положением популяций (0,80-0,95). Но очень высокая корреляция обнаружена и между генетикой и лингвистикой (0,74-0,78). Поскольку лингвистические показатели сами по себе высоко коррелируют с географией, авторы рассмотрели частную корреляцию, чтобы различить прямое и непрямое влияние географии на две другие системы. При исключении географического фактора, частная корреляция с лингвистикой стала намного ниже (0,3 для мтДНК и 0,2 для остальных двух систем), в то время как для всех трех генетических систем корреляция с географией при исключении фактора лингвистики, осталась большой (0,5 для мтДНК и 0,8 для остальных двух систем). Это указывает на то, что связь с географическим фактором – основная, а высокая связь с лингвистикой часто определяется тем, что народы, говорящие на родственных языках, являются и географическими соседями.
Два субстрата в славянских генофондах
Генетики считают, что, распространяясь по Европе, славяне ассимилировали местные популяции, которые жили на данных территориях в дославянские времена. Это тот генетический субстрат, которые они впитали в себя, и этот субстрат различается на разных территориях. Итоги работы позволили выделить два основных субстрата. «Центрально-восточноевропейский субстрат» приняли в себя западные и восточные славяне (на спектре предковых компонентов он выражается синим цветом, а в данных по Y-хромосоме эти популяции несут высокие частоты гаплогруппы R1a). Другой, «южно-восточноевропейский субстрат», впитали в себя южные славяне (это голубой цвет в спектре предковых компонентов, а особенностью Y-хромосомного генофонда являются высокие частоты гаплогруппы I2a).
В пользу этого вывода о важности субстрата в формировании генофонда славян говорят три аргумента.
Во-первых, тот факт, что объединенная группа западных и восточных славян имеет меньшее число общих фрагментов генома с южными славянами, чем с популяциями северо-восточной Европы, включая балтские и финно-угорские народы. Особая генетическая близость финно-угров с балтами видна и на графиках главных компонент, и на графиках многомерного шкалирования. А как раз народы балтской и финно-угорской языковых групп и были расселены на той части Восточно-европейской равнины, которая потом вошла в ареал славян.
Во-вторых, тест AMOVA также указывает на важную роль субстрата, поскольку генетическое разнообразие между разными ветвями славян намного превышает разнообразие внутри ветвей; такая картина и должна была сформироваться, если восточные и южные ветви славян ассимилировали генетически различные популяции.
В-третьих, преобладающая роль географии в формировании генофонда славян говорит о том же. Ведь если бы включения субстрата не было, то общее происхождение, фиксированное в языке, не могло не сказаться и на сходстве генофондов, даже когда какие-то группы славян мигрировали на далекое расстояние от своих родственников. Но такой роли лингвистического родства выявлено не было. И напротив: генетическое сходство между дославянскими популяциями, жившими на территории половины Европы, должно было быть примерно пропорционально географическим расстояниям между ними, но никак не связанным с языковым родством между славянскими группами, которые потом пришли на эти земли. Тогда, если в современных славянских генофондах преобладает субстрат, то и сходство этих генофондов должно следовать географическим расстояниям. Что и было выявлено.
Синтез данных по разным генетическим и не генетическим системам.
В генетических работах на каждом шагу встречается слово «анализ», и очень редко — «синтез». Здесь же «синтез» вынесен даже в название работы. Что это значит?
Уже говорилось о том, что это исследование уникально тем, что чуть ли не впервые для большой группы родственных народов изучен каждый народ, причем изучен по всем трем современным генетическим системам, да вдобавок количественно оценено лингвистическое родство между ними. И это позволяет на примере славян посмотреть, как связаны между собой три разные генетические системы, лингвистика и география – и синтезировать эти разнородные данные в общие выводы. Это тем более важно, что сравнение генетических и лингвистических реконструкций с географией имеет давнюю традицию в популяционной генетике.
Корреляции всех пяти систем (трех генетических, лингвистической и географической) друг с другом показаны на рисунке. Бросается в глаза очень высокое сходство всех пяти систем: ни один из коэффициентов корреляции не опускается ниже 0,68 — то есть фактически 0,7, что считается в популяционной генетике очень высокой корреляцией. А самые высокие коэффициенты достигают максимально возможного потолка (корреляция 0,95). Особенное соответствие отмечаются для Y-хромосомных и аутосомных маркеров и географического положения. Можно сказать, что эти три характеристики балто-славянских популяций образуют взаимосвязанную триаду (коэффициенты корреляции выше 0,9, темно-рыжий цвет на рисунке).
Соответствие результатов по разным системам признаков подтверждает надежность таких результатов. Оно также указывает на перспективность так называемого полисистемного подхода. Этот подход состоит, во-первых, в параллельном анализе разных систем признаков; во-вторых, в безусловном доверии лишь тем закономерностям, которые выявляются не по какой-то одной системе, а по большинству систем; в-третьих, во внимательном рассмотрении случаев, когда какая-то система выбивается из общего паттерна. В данной статье авторы широко пользовались полисистемным подходом. Формулируя утверждения о генетическом сходстве или различии тех или иных народов, авторы каждый раз проверяли, подтверждаются ли они по большинству использованных систем. А то, что из общего паттерна выбивается лингвистика, послужило одним из аргументов в пользу гипотезы преобладания субстрата.
Такой паттерн почти полного совпадения трех генетических систем друг с другом, их совпадения и с географией, но лишь частичного сходства с лингвистикой может служить маяком и для будущих исследований генофондов других регионов мира. В то же время этот паттерн не универсален для всего мира: для популяций с контрастным происхождением мужской и женской частей популяции данные по Y-хромосоме и мтДНК могут сильно различаться (что показано, например, в статье Quintano-Murci et al., 2008), а для популяций, в которых процессы постепенного роста и дробления преобладали над метисацией, генетика может быть больше скоррелирована с лингвистикой, чем с географией (что показано, например, в статье Balanovsky et al., 2011).

История славянских генофондов: итоги исследования
Прежде всего, это преобладание в славянских популяциях дославянского субстрата — двух ассимилированных ими генетических компонентов – восточноевропейского для западных и восточных славян и южноевропейского для южных славян. (Слишком длинные названия «центрально-восточноевропейский» и» южно-восточноевропейский» для краткости удобнее называть восточноевропейский и южноевропейский, помня при этом, что на Западную Европу они не простираются, а находятся в восточной ее половине при дихотомическом делении Европы).
Но, несмотря на то, что в генофонде западных и восточных славян велик ассимилированный компонент их соседей по Восточно-Европейской равнине, эти славянские популяции формируют генетически довольно целостную группу, отличающуюся как от своих западных соседей (германоязычных популяций), так и от соседей восточных и северных (финно-угорских народов). Конечно, из этого правила можно найти пару исключений, но они сосредоточены на периферии ареала западных и восточных славян. Например, у своеобразного генофонда чехов есть определенное генетическое сходство с их немецкими соседями на западе, однако другие западно-славянские популяции (поляки и сорбы) генетически четко отделяются от своих соседей-немцев. Аналогично, на другом конце славянского ареала, северные русские имеют ярко выраженное сходство с финно-угорскими и балтскими популяциями, но такого явного сходства не наблюдается для центральных или южных русских, не говоря уже о других славянских народах.
Поэтому можно предполагать, что после того как прошел основной этап распространения славянских языков и ассимиляции дославянского субстрата, началось формирование местных особенностей генофонда. Оно протекало по-разному для разных частей обширного ареала западных и восточных славян, но изначальное родство (общий субстрат плюс общий славянский суперстрат) и, вероятно, интенсивный последующий обмен генов внутри славянского ареала, сцементировали западных и восточных славян в единую генетическую общность.
В работе высказывается осторожное предположение, что ассимилированный субстрат мог быть представлен по преимуществу балтоязычными популяциями. Действительно, археологические данные указывают на очень широкое распространение балтских групп перед началом расселения славян. Балтский субстрат у славян (правда, наряду с финно-угорским) выявляли и антропологи. Полученные в этой работе генетические данные — и на графиках генетических взаимоотношений, и по доле общих фрагментов генома — указывают, что современные балтские народы являются ближайшими генетическими соседями восточных славян. При этом балты являются и лингвистически ближайшими родственниками славян. И можно полагать, что к моменту ассимиляции их генофонд не так сильно отличался от генофонда начавших свое широкое расселение славян. Поэтому если предположить, что расселяющиеся на восток славяне ассимилировали по преимуществу балтов, это может объяснить и сходство современных славянских и балтских народов друг с другом, и их отличия от окружающих их не балто-славянских групп Европы.
Что же касается южных славян, то история их генофонда могла протекать схожим образом, хотя и независимо от западных и восточных славян. Южные славяне ассимилировали значительную часть дославянского населения Балкан, которая обладала иным генофондом, чем ассимилированное восточными и западными славянами население Восточно-Европейской равнины. Потому южнославянские популяции и обнаруживают большее сходство с неславянскими популяциями Балкан (румынами и венграми), чем с другими славянскими народами.
Источник :
Genetic heritage of the Balto-Slavic speaking populations: a synthesis of autosomal, mitochondrial and Y-chromosomal data
Alena Kushniarevich, Olga Utevska, Marina Chuhryaeva, Anastasia Agdzhoyan, Khadizhat Dibirova, Ingrida Uktverite, Märt Möls, Lejla Kovačević, Andrey Pshenichnov, Svetlana Frolova, Andrey Shanko, Ene Metspalu, Maere Reidla, Kristiina Tambets, Erika Tamm, Sergey Koshel, Valery Zaporozhchenko, Lubov Atramentova, Vaidutis Kučinskas, Oleg Davydenko, Lidya Tegako, Irina Evseeva, Michail Churnosov, Elvira Pocheshchova, Bayazit Yunusbaev, Elza Khusnutdinova, Damir Marjanović, Pavao Rudan, Siiri Rootsi, Nick Yankovsky, Phillip Endicott, Alexei Kassian, Anna Dybo, The Genographic Consortium, Chris Tyler-Smith, Elena Balanovska, Mait Metspalu, Toomas Kivisild, Richard Villems and Oleg Balanovsky
http://lenta.ru/articles/2015/09/15/balto/
Передачи на радио:
Олег Балановский в передаче «Родина слонов» (радиостанция «Говорит Москва»)
http://xn--c1acc6aafa1c.xn--p1ai/wp-content/uploads/2015_09_15_Rodina_slonov.mp3Олег Балановский в передаче «Наука в фокусе» (радиостанция «Эхо Москвы»)
http://1.cdn.echo.msk.ru/snd/2015-09-18-naukafokus-1605.mp3Олег Балановский на радиостанции «Спутник» (быв. Голос России), агентство «Россия сегодня»
http://xn--c1acc6aafa1c.xn--p1ai/wp-content/uploads/151008_interview_balanovsky_genofond_researches.mp3Передачи на телевидении:
Олег Балановский в передаче «Гамбургский счет», Общественное телевидение России (ОТР)
Двойное исследование двойного континентаДве статьи, вышедшие почти одновременно в Nature и Science, посвящены генетической реконструкции заселения Америки методами анализа полных геномов. Их выводы схожи. В статье команды Давида Райха (Nature), помимо основной миграции из Сибири, давшей начало всем коренным популяциям Америки, обнаружен – пока загадочный - «австрало-меланезийский след» у некоторых популяций южноамериканских индейцев. В статье команды Эске Виллерслева (Science) обнаружен тот же след, хотя его источник мог включать, кроме Австрало-Меланезии, еще и Восточную Азию. | Происхождение славян, версия биохимическаяПубликуем рецензию археолога и филолога, доктора исторических наук Л.С.Клейна на книгу А. А. Клёсова «Происхождение славян», опубликованную в журнале "Российский археологический ежегодник". |
Как строить деревья? Проверка на лезгинских языкахВпервые проведен полноценный тест современных филогенетических методов на лексическом материале лезгинской языковой группы. | Генетическая карта Великобритании открыла окно в прошлоеИсследователи впервые создали подробную карту генетической структуры популяций Великобритании. В генофонде современной популяции удалось увидеть отражение важнейших событий в истории заселения Британских островов. |





